


Neurología funcional: curcumina para la inflamación cerebral
Índice del contenido
Efectos antiinflamatorios de la curcumina en células microgliales
Resumen
El ácido lipoteicoico (LTA) induce moléculas neuroinflamatorias, contribuyendo a la patogénesis de enfermedades neurodegenerativas. Por tanto, la supresión de moléculas neuroinflamatorias podría desarrollarse como método terapéutico. Aunque los datos anteriores apoyan un efecto inmunomodulador de la curcumina, las vías de señalización subyacentes están en gran parte sin identificar. Aquí, investigamos las propiedades anti-neuroinflamatorias de la curcumina en células microgliales BV-2 estimuladas con LTA. Factor de necrosis tumoral de citocinas inflamatorias -? La curcumina inhibió la secreción de TNF- ?, prostaglandina E2 (PGE2) y óxido nítrico (NO) en las células microgliales inducidas por LTA. La curcumina también inhibió las NO sintasas inducibles por LTA (iNOS) y la ciclooxigenasa-2 (COX-2) Posteriormente, nuestros estudios mecanicistas revelaron que la curcumina inhibía la fosforilación inducida por LTA de la proteína quinasa activada por mitógenos (MAPK), incluida ERK, p38, Akt, y la translocación de NF-? B. Además, la curcumina indujo la hemeoxigenasa (HO) -1HO- 1 y la expresión del factor 2 (Nrf-2) relacionado con el factor eritroide 2 nuclear en las células microgliales. La inhibición de HO-1 revirtió el efecto de inhibición de HO-1 sobre los mediadores inflamatorios liberados en las células microgliales estimuladas con LTA. En conjunto, nuestros resultados sugieren que la curcumina podría ser un agente terapéutico potencial para el tratamiento de trastornos neurodegenerativos mediante la supresión de respuestas neuroinflamatorias. Palabras clave: curcumina, neuroinflamación, TLR2, HO-1, células microglialesIntroducción
La neuroinflamación crónica desempeña un papel importante en diversas enfermedades neurodegenerativas, como AD, enfermedad de Parkinson (EP), enfermedad de Huntington (HD), accidente cerebrovascular, esclerosis lateral amiotrófica (ELA) y esclerosis múltiple (EM) (Spangenberg y Green, 2017). La neuroinflamación está intercedida por la activación de la microglia, las células efectoras principales y las células inmunes residentes del SNC (Nakagawa y Chiba, 2015). Las células microgliales pueden activarse en respuesta a la muerte neuronal o al daño neuronal inducido por respuestas neuroinflamatorias o por toxinas extracelulares, como bacterias y patógenos (Larochelle et al., 2015). En la neuroinflamación, la microglia activada libera varios tipos de citocinas, quimiocinas, especies reactivas de oxígeno y especies reactivas de nitrógeno para el desarrollo y mantenimiento de respuestas inflamatorias (Moss y Bates, 2001). La producción excesiva de estos mediadores inflamatorios podría causar daño neuronal y muerte. La evidencia acumulada sugiere que el control de la activación microglial podría atenuar la gravedad de la enfermedad neurodegenerativa (Perry et al., 2010). Por lo tanto, el desarrollo de agentes anti-neuroinflamatorios para la inhibición de la activación microglial podría ser beneficioso para el tratamiento de enfermedades neurodegenerativas. Microglia expresa receptores de reconocimiento de patrones (PRR) que pueden unirse a patrones moleculares asociados a patrones (PAMP) y patrones moleculares asociados a daños (DAMP) como lipopolisacárido (LPS) y ácido lipoteicoico (LTA), respectivamente (Jack et al., 2005 ) Los TLR, una clase importante de PRR, juegan un papel crucial en la defensa del huésped al inducir respuestas inmunes innatas. Cada vez más, los estudios han indicado que el agonista de TLR2 LTA está involucrado en la patogénesis de enfermedades infecciosas del SNC y puede inducir daño neuronal (Neher et al., 2011). ¿La inhibición de la activación de TLR2 atenúa la activación de células microgliales y amiloide? acumulación en el cerebro (McDonald et al., 2016; Hossain et al., 2017). La transducción de señales a través de TLR2 está mediada por diferentes proteínas adaptadoras, incluida MyD88, que promueve la señalización aguas abajo a través de la activación de MAPK y NF-? B que conduce a la expresión de mediadores inflamatorios (Larochelle et al., 2015). Las moléculas inflamatorias y oxidativas son activadores muy potentes de Keap-Nrf2 (factor 2 relacionado con NF-E2), que induce la expresión de las enzimas de desintoxicación de Fase II para adaptarse a la condición de estrés oxidativo (Rojo et al., 2010). Por lo general, Nrf2 actúa de forma inactiva. Tras la estimulación, Nrf2 se separa de Keap1 y se transloca en el núcleo, donde se une al elemento de respuesta antioxidante (ARE) para activar la transcripción de genes antioxidantes para la citoprotección (Ma, 2013; Cho et al., 2015). Uno de los genes regulados por Nrf2 es la hemo oxigenasa-1 (HO-1), que tiene una secuencia ARE en su región promotora. Recientemente, se ha informado que HO-1 es un factor predominante en el control del estrés oxidativo y las respuestas inflamatorias en enfermedades neurodegenerativas (Schipper et al., 2009). HO-1 es la primera enzima limitante de velocidad inducible en la degradación del hemo en subproductos. HO-1 puede proporcionar neuroprotección o efecto neurotóxico debido al equilibrio entre los efectos beneficiosos y tóxicos del hem y los productos de hem (Mancuso et al., 2010). Se ha demostrado que un subproducto de HO-1, la bilirrubina, protege a las neuronas del estrés oxidativo in vivo e in vitro. La bilirrubina se puede oxidar a biliverdina eliminando los radicales peroxilo (Chen, 2014). Se ha sugerido que HO-1, biliverdina y CO tienen propiedades antiinflamatorias (Jazwa y Cuadrado, 2010). Otro estudio ha sugerido que los ratones que carecen de HO-1 eran vulnerables a los estímulos proinflamatorios y desarrollaron inflamación crónica debido a la reducción de los niveles de hierro (Chora et al., 2007). Además, un estudio reciente sugirió que la regulación al alza de las vías Nrf2 y HO-1 inhibía significativamente la reacción inflamatoria en la microglia activada (Kim et al., 2016). Nrf2 inhibió la hiperactivación microglial al suprimir p38 MAPK y la vía de señalización NF-? B (Kim BW et al., 2013). Se demostró que la eliminación de Nrf2 en ratones es hipersensible a la neuroinflamación, como lo indica un aumento en los marcadores inflamatorios iNOS, IL-6 y TNF-? (Rojo et al., 2010). En consecuencia, Nrf2 y HO-1 se han considerado objetivos terapéuticos importantes para las enfermedades neurodegenerativas (Koh y col., 2011; Zhang y col., 2014). La curcumina, el principal curcuminoide aislado de Curcuma longa L. (cúrcuma) se ha utilizado durante siglos en el sudeste asiático como remedio medicinal y como alimento (Kunnumakkara et al., 2017). La curcumina, demetoxicurcumina, bisdemetoxicurcumina, ar-turmerona,? -Turmerona y? -Turmerona son los principales compuestos bioactivos que se encuentran en C. longa En estudios farmacológicos modernos, C. Los componentes de Longa, particularmente la curcumina, han mostrado actividades farmacológicas prometedoras debido a sus efectos anti-neuroinflamatorios, neuroprotectores, quimiopreventivos, inmunomoduladores y potencialmente quimioterapéuticos (Garcia-Alloza et al., 2007; Zhou et al., 2017). Un estudio anterior mostró que la curcumina inhibió las respuestas inflamatorias inducidas por LPS en los macrófagos RAW264.7, lo que sugiere un papel potencial de la curcumina en la infección bacteriana anti-Gram-negativa (Zhou et al., 2017) y la investigación in vivo e in vitro ha demostrado que la curcumina exhibe efectos antiinflamatorios (Garcia-Alloza et al., 2007; Prakobwong et al., 2011; Parada et al., 2015; Li et al., 2016). Además, también se ha informado que la curcumina promueve el desarrollo del fenotipo microglial M2 de una manera dependiente de HO-1 y reduce la inducción de iNOS, protegiendo las células microgliales contra el estrés oxidativo (Parada et al., 2015). En el presente estudio, investigamos si la curcumina podría afectar la activación microglial inducida por LTA. El ligando TLR2 LTA es un componente importante de la pared celular de las bacterias Gram-positivas.Materiales y Métodos
Materiales
La curcumina y otros reactivos se adquirieron de Sigma (C7727,> 80%, St. Louis, MO, Estados Unidos). Protoporfirina IX (SnPP) y anticuerpos dirigidos contra HO-1 (sc-390991) - Nrf2 (sc-722), proteína de unión a TATA (TBP; sc-74595),? -Tubulina (sc-134237) y? -Actina (sc-130065) - se adquirieron de Santa Cruz Biotechnology, Inc., (Dallas, TX, Estados Unidos). Anticuerpos dirigidos contra iNOS (13120): fosforilado (p) -MAPK (9910s), MAPK (9926), proteína quinasa B (Akt; 4685), p-Akt (13038) y un kit de la vía NF-? B (9936) - se adquirieron de Cell Signaling Technology, Inc., (Danvers, MA, Estados Unidos). LTA se obtuvo de InvivoGen (tlrl-pslta, Toulouse, Francia). Además, el inhibidor de JNK (inhibidor II de JNK; 420119), el inhibidor de Akt (wortmannin; 12-338), el inhibidor de ERK (PD98059, 513000) y el inhibidor de p38 (SB230580, 559395) se compraron a EMD Millipore (Billerica, MA, Estados Unidos). ). El medio de cultivo celular, DMEM y suero fetal bovino (FBS) se adquirieron de Gibco BRL (ahora Invitrogen Corporation, Carlsbad, CA, Estados Unidos).Cultivo de células
Se adquirieron células microgliales BV-2 de ratón de ATCC. Las células se cultivaron en DMEM suplementado con 10% FBS inactivado por calor y 0.1% penicilina-estreptomicina (BioSource International, Camarillo, CA, Estados Unidos) a 37 ° C en una atmósfera humidificada de 5% CO2 y 95% aire.Ensayo de viabilidad celular
La citotoxicidad de la curcumina se evaluó mediante un ensayo colorimétrico basado en microcultivo [3- (4,5-Dimethylthiazol-2-yl) -2,5-diphenyltetrazolium bromide] (MTT). Las células se incubaron en placas de pocillos 24 a una densidad de células 5 x 105 por pocillo. La solución MTT (5 ml de 5 mg / ml) se añadió a cada pocillo (concentración final 62.5 mg / ml). Después de la incubación de 3 h a 37 ° C en 5% CO2, se retiró el sobrenadante y los cristales de formazán producidos en células viables se solubilizaron con 150 ml de dimetilsulfóxido (DMSO). Luego se leyó la absorbancia de cada pocillo a 570 nm usando un lector de microplacas (Wallac 1420; PerkinElmer, Inc., Boston, MA, Estados Unidos).Medición de la concentración de nitrito
La síntesis de NO en cultivos celulares se midió mediante el método de Griess con microplaca. Para medir el nitrito, se retiraron alícuotas de 100 µl del medio acondicionado y se incubaron con un volumen igual del reactivo de Griess [sulfanilamida al 1% / clorhidrato de N- (0.1-naftil) -etilendiamina al 1% / H2.5PO3 al 4%] a temperatura ambiente durante 10 minutos. La concentración de nitrito se determinó midiendo la absorbancia a 540 nm con un espectrofotómetro de microplacas de 96 pocillos Vmax (Molecular Devices, Menlo Park, CA, Estados Unidos). Se utilizó nitrito de sodio como estándar.Medición de TNF-? y concentración de PGE2
Las células se incubaron primero con diversas concentraciones de curcumina durante 1 hora y luego con LTA durante 16 horas. Después de 24 h de incubación, TNF-? y los niveles de PGE2 se cuantificaron en los medios de cultivo utilizando un kit de ensayo inmunoabsorbente ligado a enzimas (ELISA) (R&D Systems, Minneapolis, MN, Estados Unidos) de acuerdo con las instrucciones del fabricante.Preparación de extracto nuclear
Las células microgliales BV-2 se lavaron tres veces con PBS frío y se recogieron en 3000 µl de PBS usando centrifugación a 800 xg durante 5 min (4 ° C). Los sedimentos celulares se suspendieron en tampón A [HEPES-KOH 10 mM (pH 7.9); MgCl1.5 2 mM; KCl 10 mM; Ditiotreitol (DTT) 0.5 mM; Inhibidor de proteasa (PI) 0.2 mM] y se incubó durante 5 min en hielo. Tampón B [HEPES-KOH 10 mM (pH 7.9); MgCl1.5 2 mM; NaCl 420 mM; EDTA 0.2 mM; glicerol al 25% v / v; DTT 0.1 mM; Se añadió PI 0.2 mM] al extracto celular y se incubó en hielo durante 5 min antes de la centrifugación a 11,000 xg durante 1 min a 4ºC. Las proteínas nucleares se extrajeron con la adición de tampón de lisis completo B [HEPES-KOH 10 mM (pH 7.9); MgCl1.5 2 mM; KCl 10 mM; DTT 0.5 mM; PI 0.2 mM; Glicerina al 25% (p / v); NaCl 420 mM; EDTA 0.2 mM] durante 30 min a 4ºC con agitación ocasional. Después de la centrifugación a 11,000 xg durante 5 min a 4 ° C, los sobrenadantes se recogieron y almacenaron a -70 ° C.Análisis de Western Blot
Las células BV-2 se recogieron en un tampón de lisis helado (Triton X-1 al 100%; desoxicolato al 1%; dodecilsulfato de sodio al 0.1%). El contenido de proteínas de los lisados celulares se determinó posteriormente usando reactivo de Bradford (Kit de ensayo de proteínas Bio-Rad I5000001; Bio-Rad Laboratories, Inc., Hercules, CA, Estados Unidos). Las proteínas totales de cada muestra (50 µg) se separaron mediante SDS-PAGE al 7.5% y se transfirieron a membranas de difluoruro de polivinilideno. Después del bloqueo de los sitios de unión inespecíficos con leche desnatada al 5% a temperatura ambiente durante 30 min, las membranas se incubaron con anticuerpos primarios dirigidos contra iNOS (1: 500), p-Akt (1: 1,000), p- MAPK (1: 1,000), MAPK (1: 1,000), p-p65, p65 (1: 500), pI? B ?, I? B? (1: 1,000), HO-1 (1: 1,000), Nrf2 (1: 1,000), TBP (1: 3,000),? (1: 1,000), HO-1 (1: 1.0) y actina (1: 3,000) durante 16 ha 4 ° C. A esto le siguió la incubación con anticuerpos secundarios anti-conejo (sc-2768; 1: 5,000) o anti-ratón (sc-2371; 1: 5,000) conjugados con peroxidasa de rábano picante (Santa Cruz Biotechnology, Inc.) a temperatura ambiente durante 1 h. Se usó tubulina como control de carga para cada carril. Las proteínas se visualizaron usando un kit de detección de quimioluminiscencia mejorado (GE Healthcare, Chicago, IL, Estados Unidos). Después del lavado con PBS con Tween-20, se visualizaron las bandas de proteína usando Gel Docsed como control de carga para cada carril. Las proteínas se visualizaron utilizando un analizador Quant 350 (GE Healthcare).RT-PCR en tiempo real
El ARN total se aisló de las células usando un kit de aislamiento de ARN mini ARN spin (GE Healthcare, Uppsala, Suecia) de acuerdo con las instrucciones del fabricante. Se sintetizó ADNc a partir de 1 µg de ARN total usando Maxime RT PreMix (Takara, Gyeonggi-do, Japón) y cebadores oligo-dT15 anclados. La PCR en tiempo real se realizó utilizando un instrumento Chromo4TM (Bio-Rad) y SYBR Green Master Mix (Applied Biosystems, Foster City, CA, Estados Unidos). Las cantidades relativas de ARNm diana se determinaron usando el método de umbral comparativo (Ct) normalizando los valores de Ct de ARNm diana a los de la \ beta - actina (Ct). Las secuencias primarias utilizadas en el estudio se muestran en la Tabla? 1.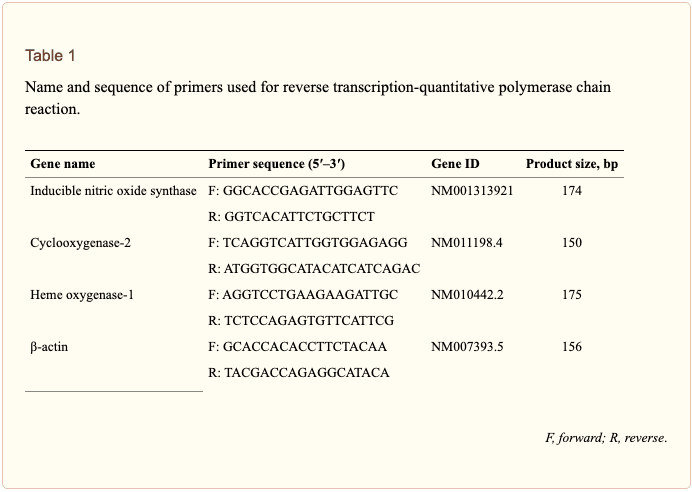
Análisis estadístico
Los datos se expresan como media (desviación estándar, DE). Cada experimento fue repetido al menos tres veces. El análisis estadístico se realizó utilizando el paquete estadístico para el software GraphPad Prism (versión 16.0) para determinar diferencias significativas. Usamos la prueba t de Student o el análisis de varianza unidireccional (ANOVA) seguido de las pruebas post hoc de Dunn para los análisis. Se consideraron estadísticamente significativos los valores de p <0.05.Resultados
La curcumina no afectó la viabilidad celular
Se llevaron a cabo experimentos de viabilidad celular para determinar si las concentraciones de curcumina utilizadas en este estudio afectaban la viabilidad de la microglía BV2. La figura? 1 muestra que la curcumina en el rango de concentración de 5-20? M, junto con o sin 5? G / ml de LTA, no produjo citotoxicidad en la microglía BV2. Por lo tanto, usamos estas concentraciones de curcumina para un estudio adicional.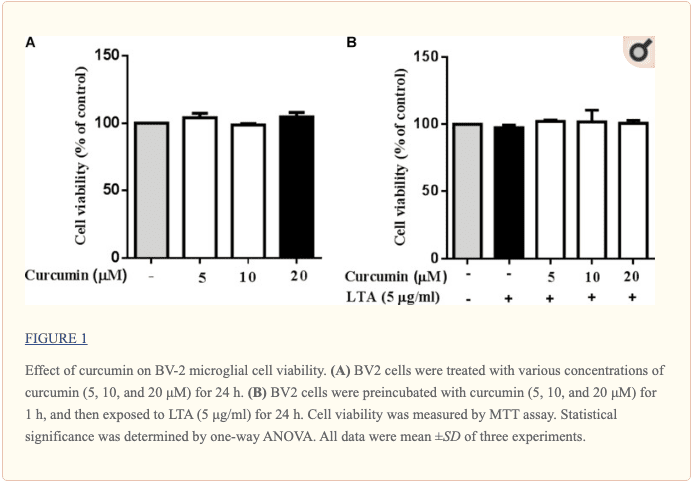
La curcumina evitó la producción de moléculas neuroinflamatorias en microglia BV2 activada por LTA
Para investigar los efectos de la curcumina sobre la secreción de citocinas inflamatorias, las células BV2 se trataron con LTA en presencia y ausencia de curcumina durante 24 h. La curcumina no se eliminó antes de la adición de LTA. Liberación de NO, PGE2 y TNF-? se redujeron de forma significativa y dependiente de la dosis con la curcumina (Figuras 2A-C). Además, LTA aumentó la expresión de ARNm de iNOS y COX-2. La incubación con curcumina suprimió la expresión de ARNm de COX-2 e iNOS en células microgliales BV2 estimuladas por LTA de una manera dependiente de la concentración (Figuras 2D, E).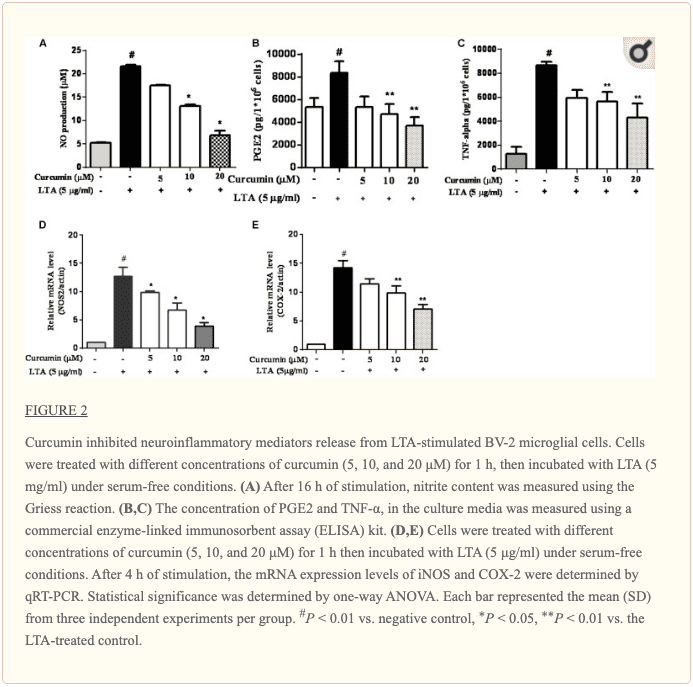
La curcumina suprimió la activación inducida por LTA de NF-? B en células microgliales BV-2
Los genes que codifican la expresión de proteínas inflamatorias en respuesta a la activación microglial estaban bajo el control de la transcripción de NF - \ kappa B. Por lo tanto, examinamos el efecto de la curcumina sobre la activación de NF - \ kappa B en células microgliales estimuladas con LTA. Los resultados mostraron que LTA indujo un aumento característico en la fosforilación de I? B ?. Después del pretratamiento con curcumina, los niveles de pI? B? se redujeron significativamente de una manera dependiente de la concentración (Figura? 3 y Figura complementaria S1). De manera consistente, la translocación nuclear de la subunidad p65 de NF - \ kappa B inducida por LTA también se atenuó mediante el tratamiento previo con curcumina. En conjunto, la curcumina probablemente atenúa la expresión de moléculas neuroinflamatorias suprimiendo la translocación nuclear y la activación de NF - \ kappa B. La cuantificación con análisis estadístico se proporcionó como datos de apoyo.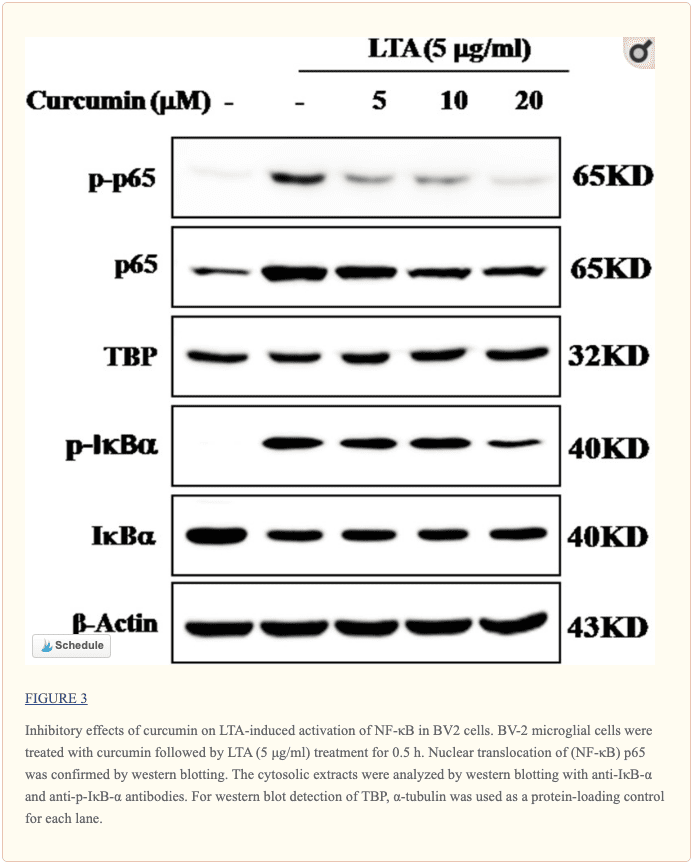
La curcumina inhibió la activación inducida por LTA de p38 y ERK MAPK en células microgliales BV-2
Aparte de NF - \ kappa B, las MAPK también son moduladores cadena arriba de moléculas neuroinflamatorias en células microgliales. Estudios previos demostraron que la curcumina antagoniza la fosforilación de MAPK inducida por LPS en microfagos (Yang et al., 2008; Kunnumakkara et al., 2017). Para investigar si la curcumina inhibe la neuroinflamación mediante la regulación de las MAPK, examinamos sus efectos sobre la fosforilación de MAPK inducida por LTA. Las células microgliales BV-2 se pretrataron con diferentes concentraciones de curcumina durante 3 horas y luego se estimularon con LTA durante 1 hora. Como se muestra en la Figura 4A y la Figura complementaria S2, la curcumina inhibió la fosforilación de ERK, p38 y Akt inducida por LTA. Sin embargo, hasta 20 µM de curcumina no afectó a la fosforilación de JNK inducida por LTA. Se ha informado que la vía MAPK media en la producción de citocinas, quimiocinas y otras moléculas neuroinflamatorias. Por lo tanto, a continuación investigamos el papel de ERK, p38, JNK y Akt en la producción de moléculas neuroinflamatorias de las células BV2 utilizando los inhibidores ERK, p38, JNK y Akt. Sin embargo, solo el inhibidor de p38 SB203580 disminuyó significativamente la liberación de NO inducida por LTA y los niveles de expresión de ARNm de iNOS (Figuras 4B, C). Aunque la curcumina no inhibió la fosforilación de JNK, el inhibidor II de JNK inhibió significativamente la liberación de NO inducida por LTA (Figura 4B). Los resultados sugieren que las vías de señalización de las MAPK están involucradas en los efectos anti-neuroinflamatorios de la curcumina en la microglía estimulada por LTA. La cuantificación con análisis estadístico se proporciona como datos de apoyo.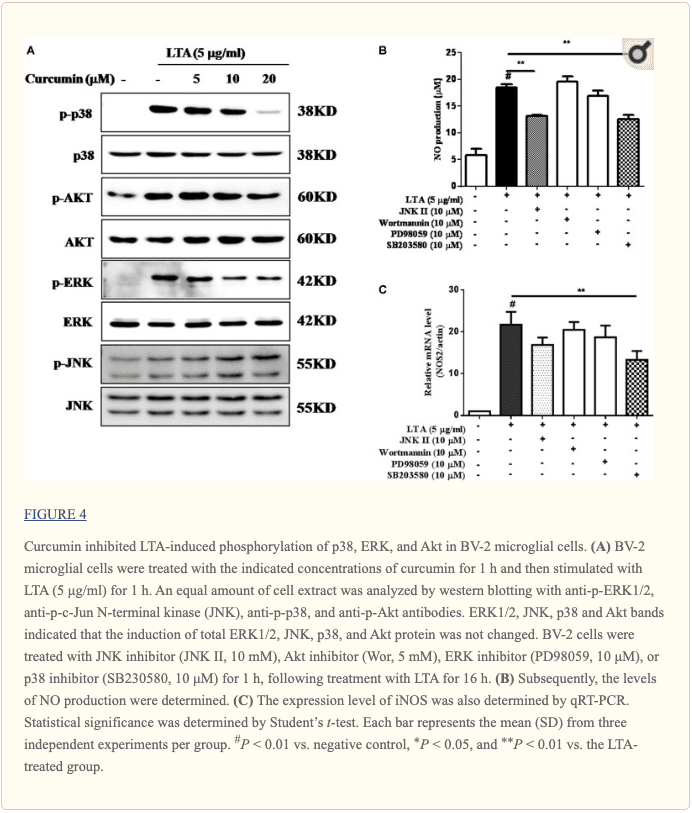
La inhibición de la señalización de HO-1 abolió el efecto inhibidor de la curcumina en las respuestas neuroinflamatorias
HO-1 actúa como un modulador antiinflamatorio y antioxidante en la microglía (Schipper et al., 2009). Los análisis de transferencia Western y RT-PCR mostraron que la curcumina regulaba al alza la expresión de HO-1 en los niveles de proteína y ARNm, como se muestra en las Figuras 5A-D y en la Figura complementaria S3. La expresión de ARNm y proteína de HO-1 aumentó al máximo en las células microgliales BV-2 tratadas con curcumina 20 µM durante 4 hy 8 h respectivamente. Además, la curcumina aumentó la translocación nuclear de Nrf2 en 1 hy prolongó su estado de translocación nuclear a 2 h (Figuras 5E, F y Figura complementaria S3). A continuación, investigamos si la HO-1 inducida por curcumina mediaba una respuesta anti-neuroinflamatoria en células microgliales BV-2 estimuladas con LTA. Tratamos células con el inhibidor de HO-1 SnPP. Luego evaluamos el efecto de la curcumina sobre el NO inducido por LTA y el TNF-? lanzamiento. El tratamiento con SnPP suprimió significativamente la inhibición mediada por la curcumina de la liberación de NO y TNF-α (Figuras 5G, H). En conjunto, estos resultados revelan que la activación de la señal HO-1 y Nrf-2 dependiente de la curcumina juega un papel crucial en la regulación a la baja de las respuestas neuroinflamatorias. La cuantificación con análisis estadístico se proporciona como datos de apoyo.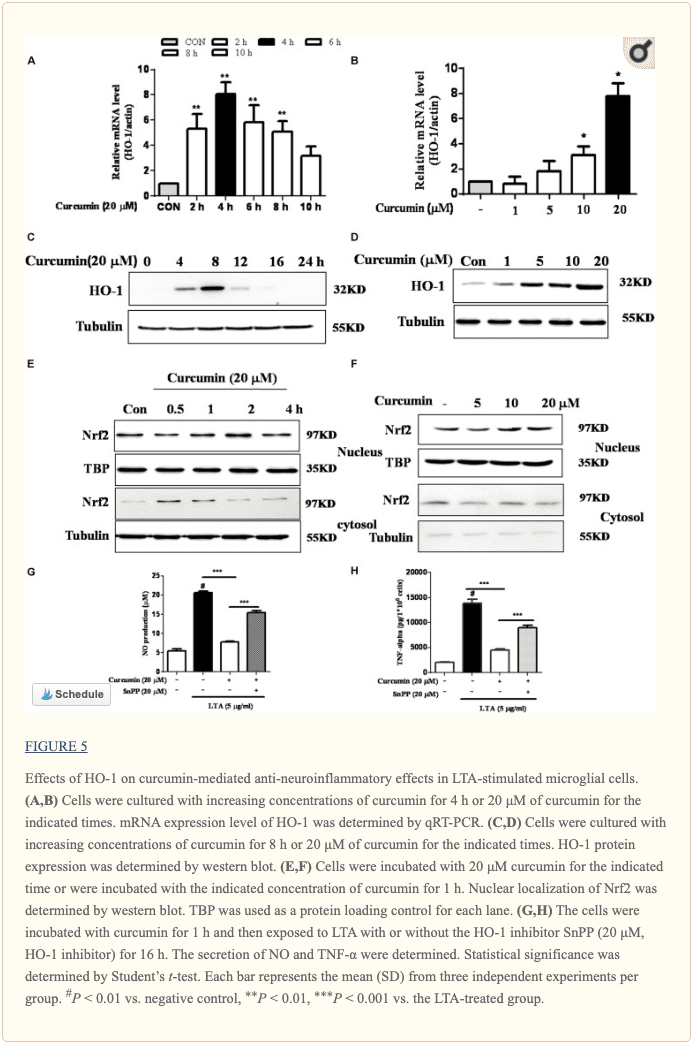
Discusión
Se ha informado que Microglia, los principales macrófagos residentes del SNC, son las principales células efectoras en la mediación de la neuroinflamación y la muerte neuronal selectiva (Perry et al., 2010). Las células microgliales aumentan la producción de moléculas neuroinflamatorias después de la exposición a activadores como LPS y LTA a través de sus receptores de superficie, TLR4 y TLR2, respectivamente (Perry y Holmes, 2014; Hossain et al., 2017). El aumento de la expresión y activación de TLR2 se asocia con la progresión de enfermedades neurodegenerativas, como la EP y la demencia (Dzamko et al., 2017). Por ejemplo, la activación de TLR2 podría regular al alza la \ beta - sinucleína en los cerebros de la EP y desempeñar funciones importantes en la patogénesis de los cerebros de la EP (Roodveldt et al., 2013). Además, Kim C. et al. (2013) también mostró que la neurodegeneración se atenuaba por la desactivación o la desactivación de TLR2 en modelos PD de roedores. Por lo tanto, el control de la activación de la microglía mediada por TLR2 y la neurotoxicidad se ha sugerido como un enfoque terapéutico importante para tratar enfermedades neurodegenerativas. Un agente potencial en este proceso podría ser la curcumina, que se ha demostrado que ejerce efectos neuroprotectores y antiinflamatorios en varios modelos experimentales (Parada et al., 2015; Li et al., 2016). La curcumina es un compuesto natural altamente lipofílico. Un estudio anterior demostró que la curcumina puede atravesar la barrera hematoencefálica y que se concentra principalmente en el hipocampo en el cerebro (Tsai et al., 2011). Algunos estudios informaron que la curcumina inhibió el daño neuronal inducido por HIV-1 gp120 y proporcionó efectos anti-neuroinflamatorios en la microglia inducida por LPS (Gong et al., 2012). Este efecto protector de la curcumina parece depender de sus acciones antiinflamatorias. La curcumina podría proteger a las neuronas contra la neurotoxicidad mediada por microglia mientras se vuelve ineficiente en condiciones de agotamiento de microglia (Park et al., 2001; Yang et al., 2008; Parada et al., 2015). Estudios similares en células periféricas también mostraron los efectos antiinflamatorios de la curcumina. Usando macrófagos murinos RAW 264.7, los estudios han demostrado que la curcumina inhibe PGE2, NO y TNF-? liberación después de la estimulación con LPS (Pae et al., 2008). Sin embargo, los efectos de la curcumina sobre la neuroinflamación inducida por TLR2 en las células microgliales no se comprenden completamente. La regulación de las vías de señalización en la microglia activada es importante para mantener la homeostasis del SNC porque las respuestas neuroinflamatorias desreguladas pueden provocar la muerte de las neuronas adyacentes a través de la liberación de moléculas inflamatorias, como citocinas, quimiocinas, NO y ROS (Perry y Holmes, 2014; Spangenberg y Green, 2017). Por ejemplo, la síntesis excesiva de NO bajo endotoxinas da como resultado la formación de especies reactivas de nitrógeno y la muerte celular neuronal (Perry et al., 2010). También se ha demostrado que PGE2 contribuye a la muerte neuronal mediante la activación de la ruta MAPK / ERK en microglia (Xia et al., 2015). En este estudio, mostramos que la curcumina inhibe la secreción de mediadores inflamatorios TNF- ?, NO y PGE2, y la expresión de iNOS y COX-2 en microglia BV2 estimulada con LTA. Además, demostramos que la curcumina atenúa estos efectos de LTA sin alterar la supervivencia celular, lo que sugiere que la curcumina es segura y podría considerarse como un posible agente terapéutico en la neuroinflamación. NF - \ kappa B es el principal factor de transcripción que juega un papel crítico en la regulación de la homeostasis redox. NF - \ kappa B se considera el regulador principal de las respuestas inflamatorias microgliales a la lesión neuronal (Acharyya et al., 2007). Estudios recientes demostraron que la activación de NF - \ kappa B controlaba la expresión de moléculas inflamatorias, como NO, PGE2 y TNF - \ beta, e IL - 1b (Acharyya et al., 2007). Por tanto, la modulación de la activación de NF - \ kappa B se considera una forma crítica de controlar la activación microglial. La activación de la ruta de señalización de NF - \ kappa B está mediada por la proteína I \ kappa B. La fosforilación de I \ kappa B da como resultado la disociación de NF - \ kappa B, que conduce a la inducción de mediadores inflamatorios. En este estudio, se demostró que la curcumina producía una inhibición dual de la fosforilación y degradación de I? B ?, así como la translocación nuclear de p65, lo que sugiere que este agente podría estabilizar NF-? B en el citoplasma microglial después de la estimulación con LTA en BV -2 células microgliales. En las células de mamíferos, las vías de señalización de MAPK, incluidas ERK, JNK y p38, contribuyen a la producción de una amplia variedad de mediadores neuroinflamatorios (Chantong et al., 2014). En este estudio actual, el pretratamiento con curcumina disminuyó la fosforilación de p38 y ERK. Además, el inhibidor p38 SB203580 redujo significativamente la secreción de NO y la expresión de ARNm del gen proinflamatorio clave, iNOS. Estos resultados sugirieron que la curcumina inició los efectos anti-neuroinflamatorios en células microgliales BV-2 estimuladas con LTA, parcialmente a través de la inhibición de la activación de p38 MAPK. La vía de señalización dependiente de PI3K / Akt promueve respuestas inflamatorias en microglia. Se ha demostrado la participación de la vía Akt en la expresión de mediadores inflamatorios en microglia a través de la activación de NF-? B en microglia (Lo et al., 2015). La curcumina suprimió el Akt fosforilado, el objetivo aguas abajo de PI3K. Sin embargo, el inhibidor de PI3K wortmannin no mostró ningún efecto inhibidor sobre la secreción de NO o la expresión de ARNm de iNOS. Tomados en conjunto, estos datos sugieren que el efecto anti-neuroinflamatorio de la curcumina se produce principalmente mediante la inhibición de la señalización de NF - \ kappa B y MAPK. También identificamos la vía intracelular que regula negativamente la expresión de la molécula inflamatoria en las células microgliales. Nrf2 es un factor de transcripción sensible a redox que regula las respuestas inflamatorias microgliales a las infecciones cerebrales. El efecto de Nrf2 se ha descrito en diferentes modelos in vivo donde la eliminación de Nrf2 en ratones aumentó la vulnerabilidad al asma o al enfisema (Ma, 2013). Además, el agonista TLR2 / TLR4 promovió respuestas inflamatorias en ratones Nrf2 KO en comparación con ratones WT (Kong et al., 2011). En el presente estudio, demostramos que la curcumina aumentó la expresión de Nrf2 y su proteína HO-1 aguas abajo. HO-1 es una molécula de señalización clave implicada en la regulación de las respuestas inflamatorias y oxidativas. El gen HO-1 tiene una secuencia ARE en su región promotora, que es un sitio de unión para el factor de transcripción Nrf2. Varios estudios han propuesto que NF-? B interrumpe la vía de señalización de Nrf-2-ARE porque muchos compuestos que indujeron la señalización de HO-1 y Nrf2 reprimieron incidentalmente la activación de NF-? B (Li et al., 2016). La expresión de HO-1 fue esencial para el efecto citoprotector específico microglial (Parada et al., 2015). Varios estudios también han demostrado una correlación inversa entre HO-1 y la secreción mediadora inflamatoria (Chora et al., 2007; Parada et al., 2015). De acuerdo, observamos que la curcumina sola indujo la expresión de HO-1 en células microgliales.Conclusión
Este estudio demostró que la curcumina tenía actividad antiinflamatoria en células microgliales estimuladas con LTA que puede inhibir la activación de NF - \ kappa B y p38 MAPK, y puede inducir la expresión de Nrf2 y HO - 1 (Figura? 6). Además, la curcumina no tiene efectos citotóxicos en las células microgliales BV-2 en su dosis antiinflamatoria. La curcumina puede tener potencial terapéutico para algunos trastornos asociados a la neuroinflamación causados por bacterias Gram-positivas.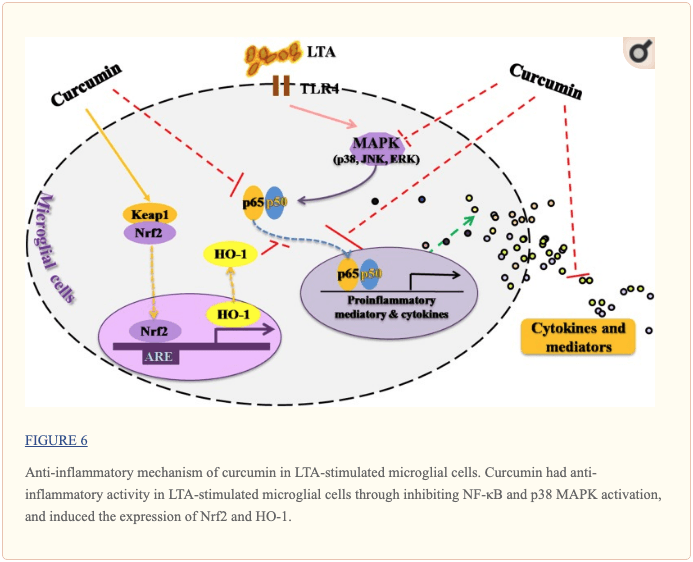

La curcumina, o cúrcuma, es un poderoso antiinflamatorio que ha demostrado tener muchos beneficios para la salud. Considerado como un antioxidante con propiedades anticancerígenas, antidepresivas y antienvejecimiento, la curcumina puede hacer mucho más que curar heridas y mejorar la memoria. Según estudios de investigación, la curcumina o la cúrcuma pueden ayudar a reducir la neuroinflamación o la inflamación cerebral. Este poderoso antiinflamatorio puede bloquear la producción de citocinas proinflamatorias y promover el bienestar general. - Dr. Alex Jiménez DC, CCST Insight
Formulario de evaluación de neurotransmisores
El siguiente formulario de evaluación de neurotransmisores se puede completar y presentar al Dr. Alex Jiménez. Los síntomas enumerados en este formulario no están destinados a ser utilizados como diagnóstico de ningún tipo de enfermedad, afección o cualquier otro tipo de problema de salud.En honor a la proclamación del gobernador Abbott, octubre es el Mes de la salud quiropráctica. Aprender más sobre la propuesta. ¿Con qué frecuencia se siente agitado, molesto y nervioso entre comidas? ¿Con qué frecuencia depende del café para mantenerse? ¿Con qué frecuencia tiene dificultad para concentrarse antes de comer? La inflamación es una respuesta importante del cuerpo humano. Es activado por el sistema inmune para protegernos contra lesiones, infecciones y / o enfermedades. Sin embargo, ¿qué sucede si hay demasiada inflamación en el cuerpo humano? Y, ¿qué sucede si hay demasiada inflamación en el cerebro? La inflamación del cerebro puede causar una variedad de problemas de salud, como ansiedad, estrés, depresión, confusión mental, fatiga e incluso letargo, entre otros síntomas comunes. Afortunadamente, existe un remedio natural que puede ayudar a reducir en gran medida la neuroinflamación y mejorar la función cerebral. Según estudios de investigación, la curcumina puede combatir la inflamación cerebral. El propósito del artículo anterior era discutir los efectos antiinflamatorios de la curcumina en la microglía y el bienestar del cerebro. El siguiente artículo ha sido referenciado por Centro Nacional de Información Biotecnológica (NCBI). El alcance de nuestra información se limita a cuestiones de salud quiropráctica, musculoesquelética y nerviosa o artículos, temas y debates sobre medicina funcional. Utilizamos protocolos funcionales de salud para tratar lesiones o trastornos del sistema musculoesquelético. Para seguir discutiendo el tema anterior, no dude en preguntarle al Dr. Alex Jiménez o contáctenos en 915-850-0900 . Comisariada por el Dr. Alex Jiménez
1 Acharyya S., Villalta SA, Bakkar N., Bupha-Intr T., Janssen PM, Carathers M., et al. (2007) La interacción de la señalización IKK / NF-kappaB en macrófagos y miofibras promueve la degeneración muscular en la distrofia muscular de Duchenne. J. Clin. Invertir. 117 889 – 901. 10.1172 / JCI30556 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
2 Chantong B., Kratschmar DV, Lister A., Odermatt A. (2014). La dibutilestaño promueve el estrés oxidativo y aumenta los mediadores inflamatorios en las células de microglia BV-2. Toxicol Letón. 230 177 – 187. 10.1016 / j.toxlet.2014.03.001 [PubMed] [CrossRef] [Google Scholar]
3 Chen J. (2014). Heme oxigenasa en neuroprotección: de mecanismos a implicaciones terapéuticas. Rev. Neurosci. 25 269 – 280. 10.1515 / revneuro-2013-0046 [PubMed] [CrossRef] [Google Scholar]
4 Cho H., Hartsock MJ, Xu Z., He M., Duh EJ (2015). El fumarato de monometilo promueve la neuroprotección dependiente de Nrf2 en la isquemia-reperfusión retiniana. J. Neuroinflamm. 12: 239. 10.1186 / s12974-015-0452-z [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
5 Chora AA, Fontoura P., Cunha A., Pais TF, Cardoso S., Ho PP, et al. (2007) Heme oxigenasa-1 y monóxido de carbono suprimen la neuroinflamación autoinmune. J. Clin. Invertir. 117 438 – 447. 10.1172 / JCI28844 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
6 Dzamko N., Gysbers A., Perera G., Bahar A., Shankar A., Gao J., et al. (2017) El receptor tipo Toll 2 aumenta en las neuronas del cerebro con enfermedad de Parkinson y puede contribuir a la patología de alfa-sinucleína. Acta Neuropathol. 133 303 – 319. 10.1007 / s00401-016-1648-8 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
7 García-Alloza M., Borrelli LA, Rozkalne A., Hyman BT, Bacskai BJ (2007). La curcumina marca la patología amiloide in vivo, altera las placas existentes y restaura parcialmente las neuritas distorsionadas en un modelo de ratón con Alzheimer. J. Neurochem. 102 1095 – 1104. 10.1111 / j.1471-4159.2007.04613.x [PubMed] [CrossRef] [Google Scholar]
8 Gong Z., Yang L., Tang H., Pan R., Xie S., Guo L., et al. (2012) Efectos protectores de la curcumina contra el virus de inmunodeficiencia humana 1 gp120 V3 lesión neuronal inducida por asa en ratas. Regeneración neuronal. Res. 7 171 – 175. 10.3969 / j.issn.1673-5374.2012.03.002 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
9 Hossain MJ, Tanasescu R., Gran B. (2017). Regulación inmune innata de la autoinmunidad en la esclerosis múltiple: concéntrese en el papel del receptor Toll-like 2. J. Neuroimmunol. 304 11 – 20. 10.1016 / j.jneuroim.2016.12.004 [PubMed] [CrossRef] [Google Scholar]
10 Jack CS, Arbor N., Manusow J., Montgrain V., Blain M., McCrea E., et al. (2005) La señalización TLR adapta las respuestas inmunes innatas en microglia y astrocitos humanos. J. Immunol. 175 4320 – 4330. 10.4049 / jimmunol.175.7.4320 [PubMed] [CrossRef] [Google Scholar]
11 Jazwa A., Cuadrado A. (2010). Dirigido a hemo oxigenasa-1 para neuroprotección y neuroinflamación en enfermedades neurodegenerativas. Curr. Objetivos de medicamentos 11 1517 – 1531. 10.2174 / 1389450111009011517 [PubMed] [CrossRef] [Google Scholar]
12 Kim BW, Koppula S., Hong SS, Jeon SB, Kwon JH, Hwang BY, et al. (2013) Regulación de la actividad de la microglía por la glaucocalixina-A: atenuación de la neuroinflamación estimulada por lipopolisacárido a través de las vías de señalización de NF-kappaB y p38 MAPK. PLoS One 8: e55792. 10.1371 / journal.pone.0055792 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
13 Kim C., Ho DH, Suk JE, You S., Michael S., Kang J., et al. (2013) La alfa-sinucleína oligomérica liberada por neurona es un agonista endógeno de TLR2 para la activación paracrina de la microglia. Nat. Commun. 4: 1562. 10.1038 / ncomms2534 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
14 Kim HJ, Kang CH, Jayasooriya RG, Dilshara MG, Lee S., Choi YH, et al. (2016) El hidrangenol inhibe la producción de óxido nítrico inducido por lipopolisacáridos en las células microgliales BV2 al suprimir la vía NF-kappaB y activar la vía HO-2 mediada por Nrf1. En t. Immunopharmacol. 35 61 – 69. 10.1016 / j.intimp.2016.03.022 [PubMed] [CrossRef] [Google Scholar]
15 Koh K., Kim J., Jang YJ, Yoon K., Cha Y., Lee HJ, et al. (2011) El factor de transcripción Nrf2 suprime la hiperactivación inducida por LPS de las células microgliales BV-2. J. Neuroimmunol. 233 160 – 167. 10.1016 / j.jneuroim.2011.01.004 [PubMed] [CrossRef] [Google Scholar]
16 Kong X., Thimmulappa R., Craciun F., Harvey C., Singh A., Kombairaju P., et al. (2011) Mejorar la vía Nrf2 mediante la interrupción de Keap1 en los leucocitos mieloides protege contra la sepsis. A.m. J. Respir. Crit. Care Med. 184 928 – 938. 10.1164 / rccm.201102-0271OC [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
17 Kunnumakkara AB, Bordoloi D., Padmavathi G., Monisha J., Roy NK, Prasad S., et al. (2017) La curcumina, el nutracéutico dorado: multitargeting para múltiples enfermedades crónicas. Br. J. Pharmacol. 174 1325 – 1348. 10.1111 / bph.13621 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
18 Larochelle A., Bellavance MA, Rivest S. (2015). Papel de la proteína adaptadora MyD88 en el preacondicionamiento y neuroprotección mediada por TLR después de la excitotoxicidad aguda. Brain Behav. Immun 46 221 – 231. 10.1016 / j.bbi.2015.02.019 [PubMed] [CrossRef] [Google Scholar]
19 Li W., Suwanwela NC, Patumraj S. (2016). La curcumina mediante la regulación negativa de NF-kB y la elevación de Nrf2 reduce el edema cerebral y la disfunción neurológica después de I / R cerebral. Microvasc. Res. 106 117 – 127. 10.1016 / j.mvr.2015.12.008 [PubMed] [CrossRef] [Google Scholar]
20 Lo JY, Kamarudin MN, Hamdi OA, Awang K., Kadir HA (2015). El curcumenol aislado de Curcuma zedoaria suprime la activación de NF-kappaB mediada por Akt y la vía de señalización de p38 MAPK en células microgliales BV-2 estimuladas con LPS. Alimento Funct. 6 3550 – 3559. 10.1039 / c5fo00607d [PubMed] [CrossRef] [Google Scholar]
21 Ma Q. (2013). Papel de nrf2 en el estrés oxidativo y la toxicidad. Annu Rev. Pharmacol. 53 401 – 426. 10.1146 / annurev-pharmtox-011112-140320 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
22 Mancuso C., Navarra P., Preziosi P. (2010). Roles de óxido nítrico, monóxido de carbono y sulfuro de hidrógeno en la regulación del eje hipotalámico-pituitario-adrenal. J. Neurochem. 113 563 – 575. 10.1111 / j.1471-4159.2010.06606.x [PubMed] [CrossRef] [Google Scholar]
23 McDonald CL, Hennessy E., Rubio-Araiz A., Keogh B., McCormack W., McGuirk P., et al. (2016) La inhibición de la activación de TLR2 atenúa la acumulación de amiloide y la activación glial en un modelo de ratón de la enfermedad de Alzheimer. Brain Behav. Immun 58 191 – 200. 10.1016 / j.bbi.2016.07.143 [PubMed] [CrossRef] [Google Scholar]
24 Moss DW, Bates TE (2001). La activación de líneas celulares microgliales murinas por lipopolisacárido e interferón-gamma causa disminuciones mediadas por NO en la función mitocondrial y celular. EUR. J. Neurosci. 13 529 – 538. 10.1046 / j.1460-9568.2001.01418.x [PubMed] [CrossRef] [Google Scholar]
25 Nakagawa Y., Chiba K. (2015). Diversidad y plasticidad de las células microgliales en trastornos psiquiátricos y neurológicos. Pharmacol El r. 154 21 – 35. 10.1016 / j.pharmthera.2015.06.010 [PubMed] [CrossRef] [Google Scholar]
26 Neher JJ, Neniskyte U., Zhao JW, Bal-Price A., Tolkovsky AM, Brown GC (2011). La inhibición de la fagocitosis microglial es suficiente para prevenir la muerte neuronal inflamatoria. J. Immunol. 186 4973 – 4983. 10.4049 / jimmunol.1003600 [PubMed] [CrossRef] [Google Scholar]
27. Pae HO, Jeong SO, Kim HS, Kim SH, Song YS, Kim SK, et al. (2008). La dimetoxicurcumina, un análogo sintético de la curcumina con mayor estabilidad metabólica, inhibe la producción de NO, la expresión inducible de NO sintasa y la activación de NF - \ kappa B en macrófagos RAW264.7 activados con LPS. Mol. Nutr. Food Res. 52 1082–1091. 10.1002 / mnfr.200700333 [PubMed] [CrossRef] [Google Académico]
28 Parada E., Buendia I., Navarro E., Avendano C., Egea J., Lopez MG (2015). La inducción microglial de HO-1 por la curcumina proporciona efectos antioxidantes, antineuroinflamatorios y glioprotectores. Mol. Nutr. Food Res. 59 1690 – 1700. 10.1002 / mnfr.201500279 [PubMed] [CrossRef] [Google Académico]
29 Park LC, Zhang H., Gibson GE (2001). El cocultivo con astrocitos o microglia protege las neuronas metabólicas. Mech Envejecimiento Dev. 123 21 – 27. 10.1016 / S0047-6374 (01) 00336-0 [PubMed] [CrossRef] [Google Scholar]
30 Perry VH, Holmes C. (2014). Cebado microglial en la enfermedad neurodegenerativa. Nat. Rev. Neurol. 10 217 – 224. 10.1038 / nrneurol.2014.38 [PubMed] [CrossRef] [Google Scholar]
31 Perry VH, Nicoll JA, Holmes C. (2010). Microglia en la enfermedad neurodegenerativa. Nat. Rev. Neurol. 6 193 – 201. 10.1038 / nrneurol.2010.17 [PubMed] [CrossRef] [Google Scholar]
32 Prakobwong S., Khoontawad J., Yongvanit P., Pairojkul C., Hiraku Y., Sithithaworn P., et al. (2011) La curcumina disminuye la colangiocarcinogénesis en los hámsters al suprimir los eventos moleculares mediados por la inflamación relacionados con la carcinogénesis multipaso. En t. J. Cáncer 129 88 – 100. 10.1002 / ijc.25656 [PubMed] [CrossRef] [Google Scholar]
33 Rojo AI, Innamorato NG, Martin-Moreno AM, De Ceballos ML, Yamamoto M., Cuadrado A. (2010). Nrf2 regula la dinámica microglial y la neuroinflamación en la enfermedad de Parkinson experimental. Glia 58 588 – 598. 10.1002 / glia.20947 [PubMed] [CrossRef] [Google Scholar]
34 Roodveldt C., Labrador-Garrido A., Gonzalez-Rey E., Lachaud CC, Guilliams T., Fernandez-Montesinos R., et al. (2013) El preacondicionamiento de microglia por alfa-sinucleína afecta fuertemente la respuesta inducida por la estimulación del receptor tipo toll (TLR). PLoS One 8: e79160. 10.1371 / journal.pone.0079160 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
35 Schipper HM, Song W., Zukor H., Hascalovici JR, Zeligman D. (2009). Heme oxigenasa-1 y neurodegeneración: expandiendo las fronteras del compromiso. J. Neurochem. 110 469 – 485. 10.1111 / j.1471-4159.2009.06160.x [PubMed] [CrossRef] [Google Scholar]
36 Spangenberg EE, KN verde (2017). Inflamación en la enfermedad de Alzheimer: lecciones aprendidas de los modelos de reducción de microglia. Brain Behav. Immun 61 1 – 11. 10.1016 / j.bbi.2016.07.003 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
37 Tsai YM, Chien CF, Lin LC, Tsai TH (2011). La curcumina y su nano formulación: la cinética de la distribución de tejidos y la penetración de la barrera hematoencefálica. En t. J. Pharm. 416 331 – 338. 10.1016 / j.ijpharm.2011.06.030 [PubMed] [CrossRef] [Google Scholar]
38 Xia Q., Hu Q., Wang H., Yang H., Gao F., Ren H., et al. (2015) La inducción de la síntesis de COX-2-PGE2 mediante la activación de la ruta MAPK / ERK contribuye a la muerte neuronal desencadenada por la microglia empobrecida por TDP-43. Celda. Muerte Dis. 6: e1702. 10.1038 / cddis.2015.69 [artículo gratuito de PMC] [PubMed] [CrossRef] [Google Scholar]
39 Yang S., Zhang D., Yang Z., Hu X., Qian S., Liu J., et al. (2008) La curcumina protege la neurona dopaminérgica contra la neurotoxicidad inducida por LPS en el cultivo primario de neurona / glía de rata. Neuroquímica Res. 33 2044 – 2053. 10.1007 / s11064-008-9675-z [PubMed] [CrossRef] [Google Scholar]
40 Zhang J., Fu B., Zhang X., Zhang L., Bai X., Zhao X., y col. (2014) Bicyclol regula al alza la expresión del factor de transcripción Nrf2 HO-1 y protege los cerebros de las ratas contra la isquemia focal. Brain Res. Toro. 100 38 – 43. 10.1016 / j.brainresbull.2013.11.001 [PubMed] [CrossRef] [Google Scholar]
41 Zhou J., Miao H., Li X., Hu Y., Sun H., Hou Y. (2017). La curcumina inhibe la inflamación placentaria para mejorar los resultados adversos del embarazo inducidos por LPS en ratones a través de la regulación positiva de Akt fosforilada. Inflamm. Res. 66 177 – 185. 10.1007 / s00011-016-1004-4 [PubMed] [CrossRef] [Google Académico]
Discusión de tema adicional: dolor crónico
El dolor repentino es una respuesta natural del sistema nervioso que ayuda a demostrar posibles lesiones. A modo de ejemplo, las señales de dolor viajan desde una región lesionada a través de los nervios y la médula espinal hasta el cerebro. El dolor generalmente es menos intenso a medida que la lesión se cura, sin embargo, el dolor crónico es diferente al tipo promedio de dolor. Con dolor crónico, el cuerpo humano continuará enviando señales de dolor al cerebro, independientemente de si la lesión se ha curado. El dolor crónico puede durar varias semanas o incluso varios años. El dolor crónico puede afectar enormemente la movilidad de un paciente y puede reducir la flexibilidad, la fuerza y la resistencia.Neural Zoomer Plus para Enfermedades Neurológicas
 El Dr. Alex Jiménez utiliza una serie de pruebas para ayudar a evaluar enfermedades neurológicas. El zoomero neuralTM Plus es un conjunto de autoanticuerpos neurológicos que ofrece reconocimiento específico de anticuerpo a antígeno. El zoomero neuronal vibranteTM Plus está diseñado para evaluar la reactividad de un individuo a los antígenos neurológicos 48 con conexiones a una variedad de enfermedades relacionadas neurológicamente. El zoomero neuronal vibranteTM Plus tiene como objetivo reducir las condiciones neurológicas al brindar a los pacientes y médicos un recurso vital para la detección temprana de riesgos y un enfoque mejorado en la prevención primaria personalizada.
El Dr. Alex Jiménez utiliza una serie de pruebas para ayudar a evaluar enfermedades neurológicas. El zoomero neuralTM Plus es un conjunto de autoanticuerpos neurológicos que ofrece reconocimiento específico de anticuerpo a antígeno. El zoomero neuronal vibranteTM Plus está diseñado para evaluar la reactividad de un individuo a los antígenos neurológicos 48 con conexiones a una variedad de enfermedades relacionadas neurológicamente. El zoomero neuronal vibranteTM Plus tiene como objetivo reducir las condiciones neurológicas al brindar a los pacientes y médicos un recurso vital para la detección temprana de riesgos y un enfoque mejorado en la prevención primaria personalizada.
Fórmulas para soporte de metilación
 De XYMOGEN Las fórmulas profesionales exclusivas están disponibles a través de profesionales de atención médica con licencia seleccionados. La venta por internet y el descuento de fórmulas XYMOGEN están estrictamente prohibidos.
Con orgullo El Dr. Alexander Jimenez hace que las fórmulas de XYMOGEN estén disponibles solo para los pacientes bajo nuestro cuidado.
Llame a nuestro consultorio para que podamos asignar una consulta médica para acceso inmediato.
Si eres paciente de Clínica médica y quiropráctica de lesiones, puedes preguntar por XYMOGEN llamando 915-850-0900.
De XYMOGEN Las fórmulas profesionales exclusivas están disponibles a través de profesionales de atención médica con licencia seleccionados. La venta por internet y el descuento de fórmulas XYMOGEN están estrictamente prohibidos.
Con orgullo El Dr. Alexander Jimenez hace que las fórmulas de XYMOGEN estén disponibles solo para los pacientes bajo nuestro cuidado.
Llame a nuestro consultorio para que podamos asignar una consulta médica para acceso inmediato.
Si eres paciente de Clínica médica y quiropráctica de lesiones, puedes preguntar por XYMOGEN llamando 915-850-0900.
Publicar descargo de responsabilidad *
Alcance de la práctica profesional *
La información aquí contenida en "Neurología funcional: curcumina para la inflamación cerebral" no pretende reemplazar una relación personal con un profesional de la salud calificado o un médico con licencia y no es un consejo médico. Lo alentamos a que tome decisiones de atención médica basadas en su investigación y asociación con un profesional de la salud calificado.
Información del blog y debates sobre el alcance
Nuestro alcance informativo se limita a la quiropráctica, musculoesquelética, medicina física, bienestar, contribuyendo etiológico alteraciones viscerosomáticas dentro de las presentaciones clínicas, la dinámica clínica del reflejo somatovisceral asociado, los complejos de subluxación, los problemas de salud delicados y/o los artículos, temas y debates de medicina funcional.
Brindamos y presentamos colaboración clínica con especialistas de diversas disciplinas. Cada especialista se rige por su ámbito de práctica profesional y su jurisdicción de licencia. Utilizamos protocolos funcionales de salud y bienestar para tratar y apoyar la atención de lesiones o trastornos del sistema musculoesquelético.
Nuestros videos, publicaciones, temas, asuntos e ideas cubren cuestiones clínicas, problemas y temas que se relacionan y respaldan directa o indirectamente nuestro ámbito de práctica clínica.*
Nuestra oficina ha intentado razonablemente proporcionar citas de apoyo y ha identificado el estudio o los estudios de investigación relevantes que respaldan nuestras publicaciones. Proporcionamos copias de los estudios de investigación de respaldo disponibles para las juntas reguladoras y el público a pedido.
Entendemos que cubrimos asuntos que requieren una explicación adicional de cómo puede ayudar en un plan de atención o protocolo de tratamiento en particular; por lo tanto, para discutir más a fondo el tema anterior, no dude en preguntar Dr. Alex Jiménez, DC, o póngase en contacto con nosotros en 915-850-0900.
Estamos aquí para ayudarlo a usted y a su familia.
Bendiciones
El Dr. Alex Jimenez corriente continua MSACP, enfermero*, CCCT, IFMCP*, CIFM*, ATN*
email: coach@elpasomedicinafuncional.com
Licenciado como Doctor en Quiropráctica (DC) en Texas & New Mexico*
Número de licencia de Texas DC TX5807, Nuevo México DC Número de licencia NM-DC2182
Licenciada como Enfermera Registrada (RN*) en Florida
Licencia de Florida N.° de licencia de RN RN9617241 (Control No. 3558029)
Estado compacto: Licencia multiestatal: Autorizado para ejercer en 40 Estados*
Matriculado actualmente: ICHS: MSN* FNP (Programa de enfermera practicante familiar)
Dr. Alex Jiménez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Mi tarjeta de presentación digital
